
Care este utilitatea unui sfert sau a unei cincimi dintr-o bătaie a aripii? Este posibil ca o insectă sau o pasăre să supraviețuiască cu o aripă care este mult mai mică decât dimensiunea normală? Și-ar putea aceasta îndeplini scopul funcționând la o capacitate mai redusă? Aceste întrebări reprezintă o provocare pentru cei care acceptă gradualismul și adaptarea darwiniană. Potrivit teoriei evoluției darwiniste, trăsăturile biologice se dezvoltă prin mici variații genetice care modifică în mod continuu structurile corporale și care sunt supuse procesului de selecție naturală. Noile trăsături sunt selectate sau eliminate în lupta pentru supraviețuire și adaptare la mediu. În acest context, o aripă proiectată pentru zbor și care nu este complet formată sau funcțională nu ar fi utilă, iar selecția naturală ar elimina-o.
O pasăre sau o insectă nu poate zbura cu o cincime de aripă sau cu o cincime din bătaia unei aripi. Cu toate acestea, evoluționiștii își doresc să ne convingă că aceste structuri au trecut printr-o serie de etape intermediare până la atingerea dimensiunii complete, în decursul unei perioade extinse de evoluție. Această afirmație ridică două întrebări: 1. Există vreun caz de forme intermediare în registrul fosil al păsărilor și insectelor? 2. Care ar fi utilitatea unei aripi incomplet formate?
În conformitate cu modelul darwinian, mutațiile benefice, capabile să ducă la formarea unor aripi complet dezvoltate, s-au acumulat pe parcursul a milioane de ani prin etape treptate, evitând constrângerile selecției naturale, care au tendința de a elimina trăsăturile inutile, ceea ce a condus în cele din urmă la apariția structurilor de aripi pe care le vedem acum la păsări și insecte. Dacă ar fi așa, ne-am aștepta să vedem mai multe forme intermediare în arhiva fosilă. Cu toate acestea, nu este cazul.
Aripile in Registrul Fosilelor
Păsările apar în registrul fosilelor cu aripi complet formate pentru zbor. Presupușii intermediari – Dromeosauridele și Archaeropteryx – prezintă un amestec între anumite trăsături de reptilă și anumite trăsături de păsări. De altfel, presupușii intermediari nici măcar nu sunt cei mai buni candidați pentru presupusele forme intermediare, deoarece prezintă ceea ce paleontologii numesc morfologie mozaicată, în care presupusa linie genealogică nu prezintă o secvență de schimbări graduale, ci numai discontinuități, inversări și trăsături ieșite din secvență.[1]
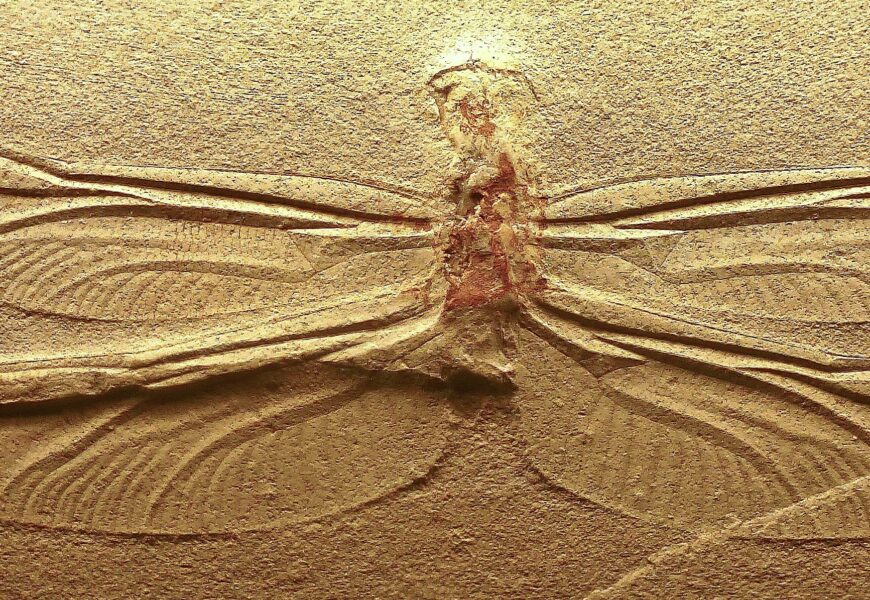
Aripile insectelor prezintă o trăsătură complexă, care apare brusc în registrul fosilelor, fără niciun strămoș cu trăsături intermediare sau de tranziție. În cadrul cronologiei evolutive, insectele cu aripi, capabile să zboare, apar în registrul fosilelor în Carboniferul superior, în urmă cu aproximativ 320 de milioane de ani. Fosilele de insecte fără aripi apar în Silurian, cu aproximativ 80 de milioane de ani înainte, din câte se presupune.[2] Nu au fost găsite fosile care să arate stadii intermediare în evoluția de la forme fără aripi la insecte cu aripi. Aceast lucru ridică următoarea întrebare: cum a fost posibil ca în cei 80 de milioane de ani de presupusă evoluție să nu fi rămas nicio dovadă a unor forme de tranziție?
Modelarea originii aripilor la insecte
Au fost propuse mai multe ipoteze privind originea aripilor la insecte. În 1985, Kingsolver și Koehl[3] au analizat două ipoteze care includeau factori de adaptare ce ar fi putut favoriza tranziția de la insecte care nu zboară la insecte cu aripi capabile să zboare. Acești factori de adaptare au inclus aerodinamica (planare și parașutare) și termodinamica. Kingsolver și Koehl au indicat că, la început, aripile ar fi putut funcționa ca profile aero-dinamice pentru planare în stare constantă, ceea ce le-ar fi permis insectelor să sară de la sol sau din copaci și astfel să evite prădătorii. Alternativ, este posibil ca primele aripi să se fi dezvoltat ca parașute care încetineau viteza de coborâre a unei insecte aflate în cădere sau s-ar fi putut dezvolta ca structuri de termoreglare pentru a crește temperatura corpului prin absorbția radiațiilor, favorizând astfel o activitate mai viguroasă. După ce au realizat modele pentru diferite forme și mărimi de insecte și după ce au făcut diverse experimente cu modele artificiale în tuneluri de vânt cu circuit deschis și cu jet, au concluzionat că niciunul dintre acești factori nu ar putea fi considerat un element determinant pentru evoluția aripilor de insecte. Există mai multe elemente incerte, cum ar fi condițiile ecologice din trecut, care ar fi putut afecta nevoile insectelor, și totodată condițiile climatice necunoscute din perioada Devoniană (a fost climatul rece cel care a determinat dezvoltarea termoreglării?).
Kingsolver și Koehl au sugerat că aripile insectelor au evoluat pornind de la structuri mici, rudimentare, care au fost preadaptate (pre-programate) în ceea ce privește zborul. Cu alte cuvinte, evoluția a fost direcționată și a avut un scop pe termen lung, dezvoltând unele structuri timpurii care aveau să servească mai târziu ca organe de zbor, dar care între timp serveau altor scopuri. Această idee vine cumva în contradicție cu presupunerea că mutațiile și selecția naturală sunt forțe ale naturii neghidate, oarbe și fără scop.[4] În plus, aceasta este o explicație nesatisfăcătoare și, într-adevăr, nu rezolvă problema lipsei de dovezi pentru evoluția aripilor la insecte. Lipsa de dovezi pentru o idee nu se rezolvă cu invocarea unei alte idei pentru care, de asemenea, nu există dovezi care să o susțină.
Au evoluat aripile ca structuri ,,preadaptate”?
Ideea de preadaptare sau de pre-programare se regăsește și în alte ipoteze privind originea aripilor la insecte. Luați în considerare, de exemplu, ,,teoria aripioarelor”, ideea sugerată de Bradley[5] și revizuită de Kukalova-Peck,[6] conform căreia pterigoții, un grup de artropode antice, în principal terestre,[7] au devenit înotători și, în timp, s-au adaptat din ce în ce mai mult pentru a se deplasa în apă. Potrivit acestei teorii, artropodele acvatice primitive prezentau mici proeminențe pe partea toracică, sau protoaripi, care ulterior au evoluat în aripi. Bradley a sugerat că ,,micile aripi care ar fi avut un efect mic sau chiar inexistent într-un posibil zbor ar fi putut fi extrem de utile ca aripioare și organe de propulsie în apă. În procesul utilizării aripilor primitive ca înotătoare, articulația și musculatura de zbor au început să se dezvolte.”[8] El a sugerat, de asemenea, că pterigoții erau probabil capabili să părăsească apa pentru împerechere și răspândire.
Ce arată insectele ,,primitive”?
În sprijinul acestei teorii, Bradley a prezentat fosile ca dovezi: toate nimfele paleozoice primitive[9] aveau aripioare articulate și mobile pe tot parcursul dezvoltării lor ontogenetice.[10] Deși nu prezintă nicio dovadă în acest sens, el afirmă că aceste aripi au fost folosite pentru o funcție de propulsie. Potrivit lui Kukalova-Peck, este ,,destul de evident că utilizarea aripilor la unele nimfe paleozoice în condiții aeriene era deja în scădere. Astfel, aripile tuturor gândacilor tineri tereștri erau complet lipsite de abilitatea de a zbura. Unele aripi de nimfă la Palaeodictyoptera, după cum reiese din forma și poziția lor generală, erau în curs de a-și pierde rapid capacitatea de funcționare. În același timp, nimfele de efemeride acvatice își foloseau aripile pentru a se deplasa în apă.”[11] Aici autorul spune că în Paleozoic existau nimfe terestre cu aripi, care își pierdeau treptat funcționalitatea. Ca urmare a acestei evoluții, unele insecte terestre nu au mai zburat și, în același timp, unele artropode acvatice au început să folosească lobii corpului pentru a se mișca sub apă, care mai târziu aveau să se transforme în aripi. Întregul scenariu este fictiv și contradictoriu: inițial, apar aripi la insectele terestre; apoi, acestea se micșorează sau chiar dispar; în același timp, se dezvoltă aripi (pentru propulsie) la artropodele acvatice; și, în cele din urmă, există o tranziție treptată de la mediu acvatic la terestru, cu o nouă dezvoltare a aripilor la insectele de pe uscat. Există dovezi pentru fiecare dintre etapele presupuse? Nu. Singurele lucruri pe care le avem la dispoziție sunt fosilele de insecte înaripate și fosilele de insecte fără aripi, plus alte artropode. Dar fosilele în sine nu demonstrează evoluția. Ele arată doar ceea ce a existat: artropode cu aripi și artropode fără aripi, prezente atât în mediul acvatic, cât și în cel terestru.
Mai mult, modelul Kulakova-Peck se confruntă cu o problemă foarte importantă: pterigoții, presupusul strămoș al insectelor înaripate, apar pentru prima dată în arhiva fosilă în straturi de rocă aflate deasupra straturilor cu insecte înaripate.[12] Așadar, ele nu se succed. Presupusul strămoș apare după formele diversificate care sunt considerate a fi descendente, aflându-se într-un scenariu similar cu cel al păsărilor. În acest sens, pentru a evita necesitatea de a explica de două ori originea aripilor, întâi pentru insectele terestre și apoi pentru artropodele acvatice, evoluționiștii susțin că evoluția pterigoților a avut loc într-o etapă mai timpurie, generând forme înaripate ale insectelor și menținându-se, de asemenea, ca forme acvatice fără aripi. A găsit cineva ca dovezi fosile pentru astfel de speculații? Nu s-a găsit nicio dovadă în registrul fosilelor pentru presupusele tranziții de la uscat la apă și de la apă la uscat, ci doar presupuneri derivate din teoria evoluționistă.
Evoluția vrea să explice totul
Ideea că zborul s-a dezvoltat la insectele terestre pornind de la strămoși acvatici este lipsită de dovezi fosile care să o susțină, cu excepția unor exemple izolate de specimene cu aripi și fără aripi. Adăugând și mai mult la raționamentul defectuos și urmând strategia evoluționistă obișnuită, Kukalova-Peck spune că ,,ipoteza lui Bradley (teoria ,,aripioarelor”), chiar dacă explică în mod eronat originea aripii în sine, pare să fie cumva conformă cu dovezile fosile în ceea ce privește originea zborului”.[13] Am observat că înregistrările fosile nu prezintă evoluția, ele doar reflectă ceea ce exista acolo. Kukalova-Peck consideră că înregistrările fosilelor ar putea susține o ipoteză, care, în același timp ar putea fi incorectă din alte considerente. Se ridică următoarea întrebare: care este rolul registrului fosilelor? Nu ar trebui să indice dovezi în favoarea sau împotriva unor idei probabile? Acest lucru este tipic pentru raționamentul materialist: ipotezele, presupunerile sau modelele nu sunt niciodată puse la îndoială, deoarece ei știu că evoluția este oricum adevărată, chiar dacă datele semnalează contrariul.
S-au făcut și alte speculații cu privire la modul în care au apărut aripile la insecte, una dintre ele fiind ideea că aripile s-ar fi dezvoltat inițial ca structuri toracice folosite în ritualuri de împerechere la masculii insectelor[14] sau că segmentele toracice și abdominale ale nimfelor acvatice s-au dezvoltat ca structuri folosite pentru ventilație și/sau schimbul de gaze.[15] Din nou, nu există nicio dovadă pentru aceste idei. Ele derivă din necesitatea de a explica modul în care insectele cu aripi au apărut în cadrul unei paradigme care presupune evoluția lor dintr-un strămoș comun.
Evoluția aripilor care nu zboară
Registrul fosilelor la insecte arată apariția bruscă atât a insectelor care nu pot zbura, cât și a celor care pot zbura, fără nicio formă de tranziție. Registrul fosil indică existența unor specii care au o complexitate ridicată, atât în ceea ce privește anatomia, cât și fiziologia, și care sunt adaptate complet la medii terestre sau acvatice. Cu toate acestea, nu se găsesc forme intermediare, nici din punct de vedere al aspectului morfologic, nici al habitatului ecologic. În plus, chiar dacă ar exista astfel de forme, cum am putea ști că sunt forme intermediare de evoluție și nu doar forme ecologice diferite, fără legături între ele?
Kinsolver și Koehl au identificat o chestiune importantă asociată cu registrele fosilelor în ceea ce privește evoluția aripilor insectelor. Ei au formulat întrebarea: „Ce caracteristici au avut membrii grupului ancestral al pterigoților (posibilii strămoși ai insectelor cu aripi) înainte de apariția completă a aripilor și a capacității de zbor?”[16]. Mai pe scurt, cum arătau strămoșii insectelor cu aripi? După cum menționează autorul, paleontologii speculează că, în cazul păsărilor, unele specii, cum ar fi Archaeopteryx, ar putea oferi indicii importante pentru înțelegerea evoluției. Cu toate acestea, absența totală a fosilelor de insecte de tranziție între perioada Devonianului timpuriu și sfârșitul Carboniferului reprezintă o provocare semnificativă pentru cercetările care încearcă să facă analogii cu privire la evoluția aripilor și a zborului la insecte.[17] Aici presupunem că atât ipoteza evoluționistă a formelor de tranziție, cât și timpul implicat sunt greșite.
Articol poate fi găsit accesând următorul link: https://www.grisda.org/the-perfect-wing-stroke-1#_edn11
Articol publicat la data de 3 decembrie 2013, de către Raul Esperante, Geoscience Research Institute
Articol tradus de George Rad
[1]Pentru o recenzie, a se vedea T. Standish, Aves fósiles, Ciencia de los Orígenes 67 (mai 2004): 1-5; R. Esperante, ¿A dónde ha volado la secuencia evolutiva de los reptiles a las aves? Ciencia de los Orígenes, 68 (toamna 2004:8-9).
[2] Engel, M. and Grimaldi, D., 2004. New light shed on the oldest insect. Nature, 427: 627-630.
[3] Kingsolver, J.G. and Koehl, M. 1985. Aerodynamics, thermoregulation, and the evolution of insect wings: differential scaling and evolutionary change. Evolution, 39(3):488-504.
[4] Douglas J. Futyuma, în cartea sa Evolution (Sinauer Associates, Sunderland, Connecticut, 2009), p. 282, afirmă că “[Alternativa lui Darwin] la designul inteligent a fost designul prin procesul complet lipsit de sens al selecției naturale, conform căruia organismele care posedă variații ce îmbunătățesc supraviețuirea sau reproducerea, le înlocuiesc pe cele mai puțin înzestrate, care, prin urmare, supraviețuiesc sau se reproduc în mai mică măsură. Acest proces nu poate avea un scop, așa cum nici eroziunea nu are scopul de a forma canioane, deoarece viitorul nu poate provoca evenimente materiale în prezent. Astfel, conceptele de ținte sau scopuri nu-și au locul în biologie (sau în oricare altă știință naturală), cu excepția studiilor privind comportamentul uman.”
[5] Bradley, J. C. 1942 . The origin and significance of metamorphosis and wings among insects. Proc. VIII. Pan Amer. Sci. Congr., Biol. Sec., 3:303-309.
[6] Kukalova‐Peck, J. 1978. Origin and evolution of insect wings and their relation to metamorphosis, as documented by the fossil record. Journal of Morphology, 156:53-125.
[7] Unul dintre cei mai comuni reprezentanți din arhivele fosile este Pterygotus, care a fost un gen antic de artropode marine mari, printre care se numără și scorpionii de mare. Aceștia aveau patru perechi de picioare pentru mers, o pereche de palete de înot și o pereche de chelae (clești) mari pentru capturarea prăzii. Acestea erau organisme marine complet dezvoltate și adaptate, care nu prezintă nicio dovadă de tranziție de la sau la orice altă formă animală.
[8] Citat din Kukalova-Peck, p. 69.
[9] Nimfele sunt forme ce nu au ajuns la maturitate ale unor nevertebrate, inclusiv ale insectelor, care suferă o metamorfoză treptată înainte de a ajunge la stadiul de adult.
[10] Dezvoltarea ontogenetică (sau ontogeneza) reprezintă procesul de dezvoltare a unui individ de la momentul conceperii, trecând prin faza de embrion, prin stadiul de făt, apoi prin procesul nașterii și prin cel al creșterii până la stadiul de adult.
[11] Kukalova-Peck, p. 70.
[12] Kingsolver, J.G. și Koehl, M. (1994) Selective factors in the evolution of insect wings. Annual Review of Entomology, 39, 425-451. Mai precis, pe scara timpului geologic, insectele pterigote (acele artropode acvatice cu presupuse protoaripi) au apărut după insecte precum paleopterus și neopterus, care aveau deja aripi complet formate.
[13] Kukalova-Peck, p. 70, emphasis added.
[14] Alexander, R.D. and Brown Jr, W. (1962) Mating behavior and the origin of insect wings. Occasional Papers of the Museum of Zoology, University of Michigan, 628:1-19.
[15] Kingsolver and Koehl, 1994.
[16] Kingsolver and Koehl, 1994, p. 429. Emphasis added.
[17] Ibid. Emphasis added.
